Transkripsjon i bakterier
© Anders Skovly 2024
Både transkripsjon og oversetting utføres noe forskjellig hos bakterier i forhold til eukaryoter. Denne artikkelen handler om transkripsjon i bakterier.
Transkripsjonsprosessen
Transkripsjon er en slags kopiering av en basesekvens fra DNA til RNA. Kopieringen er ikke eksakt: mens adeniner, cytosiner og guaniner kopieres "en-til-en", så kopieres thyminer i DNA til de lignende uracil i RNA. Hovedkomponenten i transkripsjonsprosessen er proteinet RNA-polymerase (RNApol). I RNApol finnes et Produkt-sted og et Addisjon-sted (P-sted og A-sted), som hver kan binde en ribose-nukleotid.
Transkripsjon av et gen begynner med at RNApol binder til DNA nær starten av genet, ved en basesekvens kalt promoter. Deretter brytes en rekke basepar i promoteren, og dobbelheliksens to DNA skiller seg og danner en transkripsjonsboble. En av de skilte DNA vil brukes direkte i syntesen av RNA, denne DNA kalles templat. Den andre DNA kalles nontemplat. Hvilken av de to DNA som blir templat varierer fra gen til gen og avgjøres av promoteren.
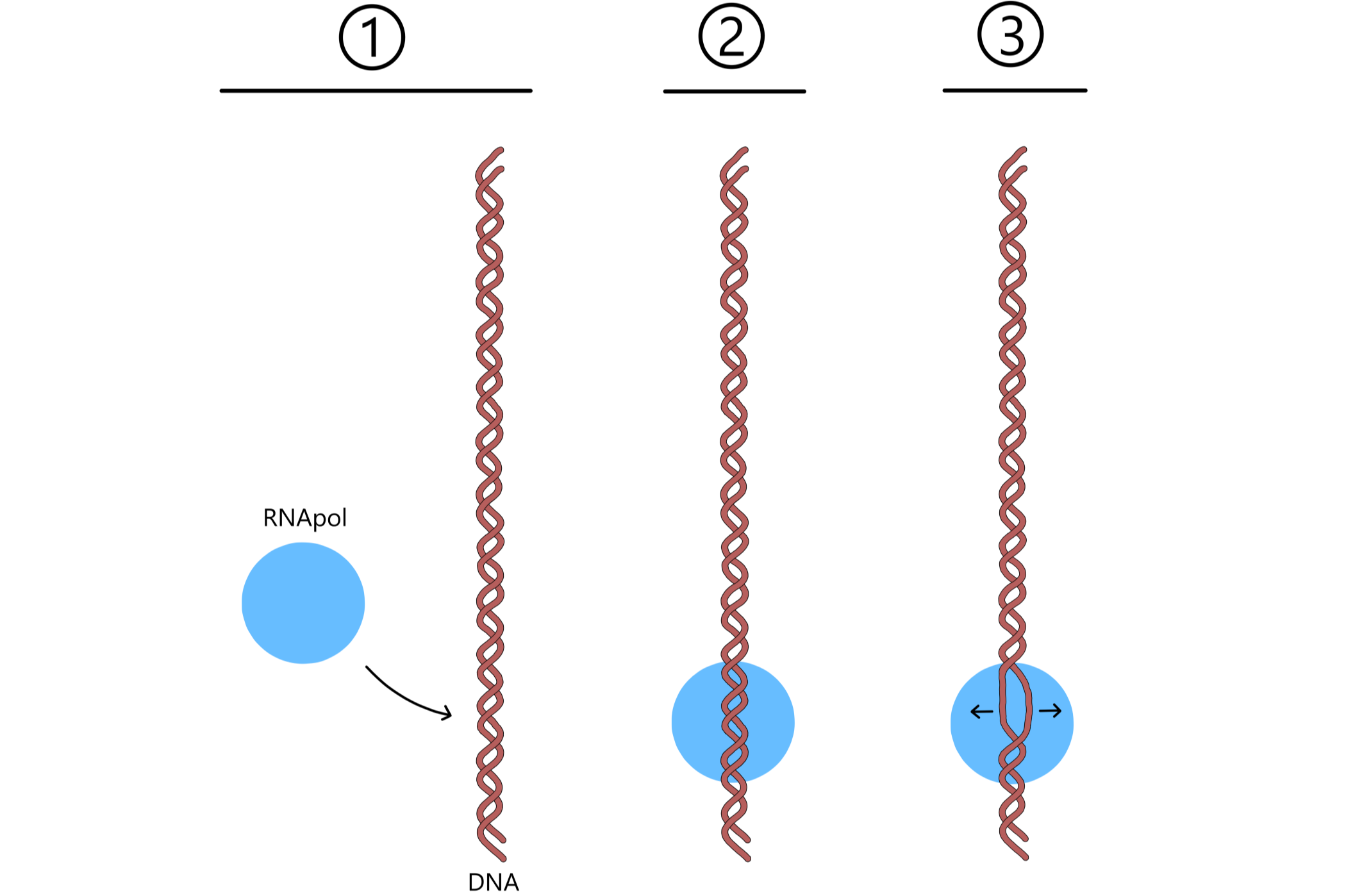
Templatet posisjoneres på en slik måte at en templat-base vil være nær P-stedet, og en "nabo" templat-base vil være nær A-stedet. Disse basene vil kalles første og andre base i templatet. En ribose-nukleotid entrer P-stedet, og dersom nukleotiden er komplementær til templatets første base vil de to danne et basepar. På samme måte vil også en ribose-nukleotid entre A-stedet, og dersom denne er komplementær til templatets andre base vil de to danne et basepar.
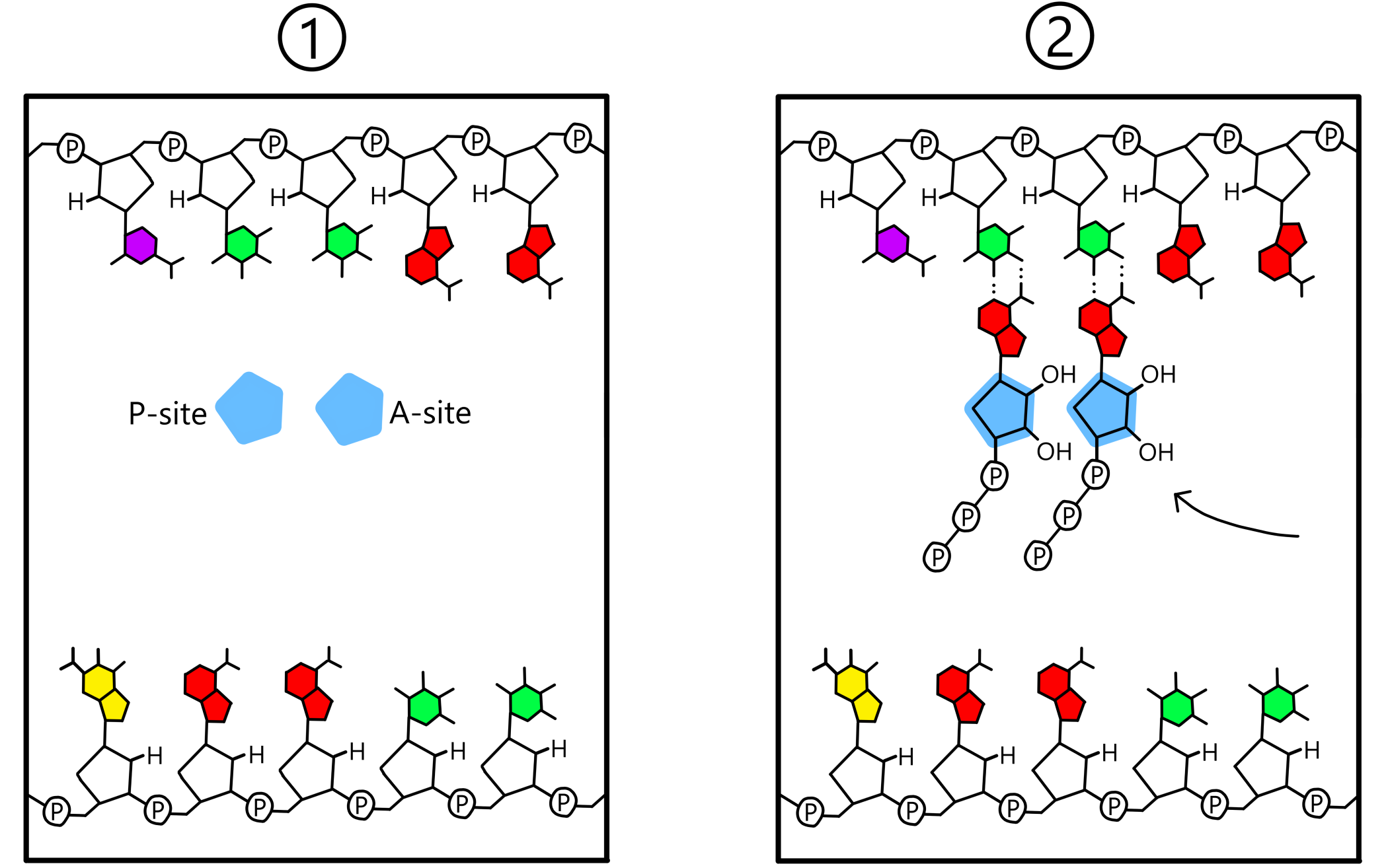
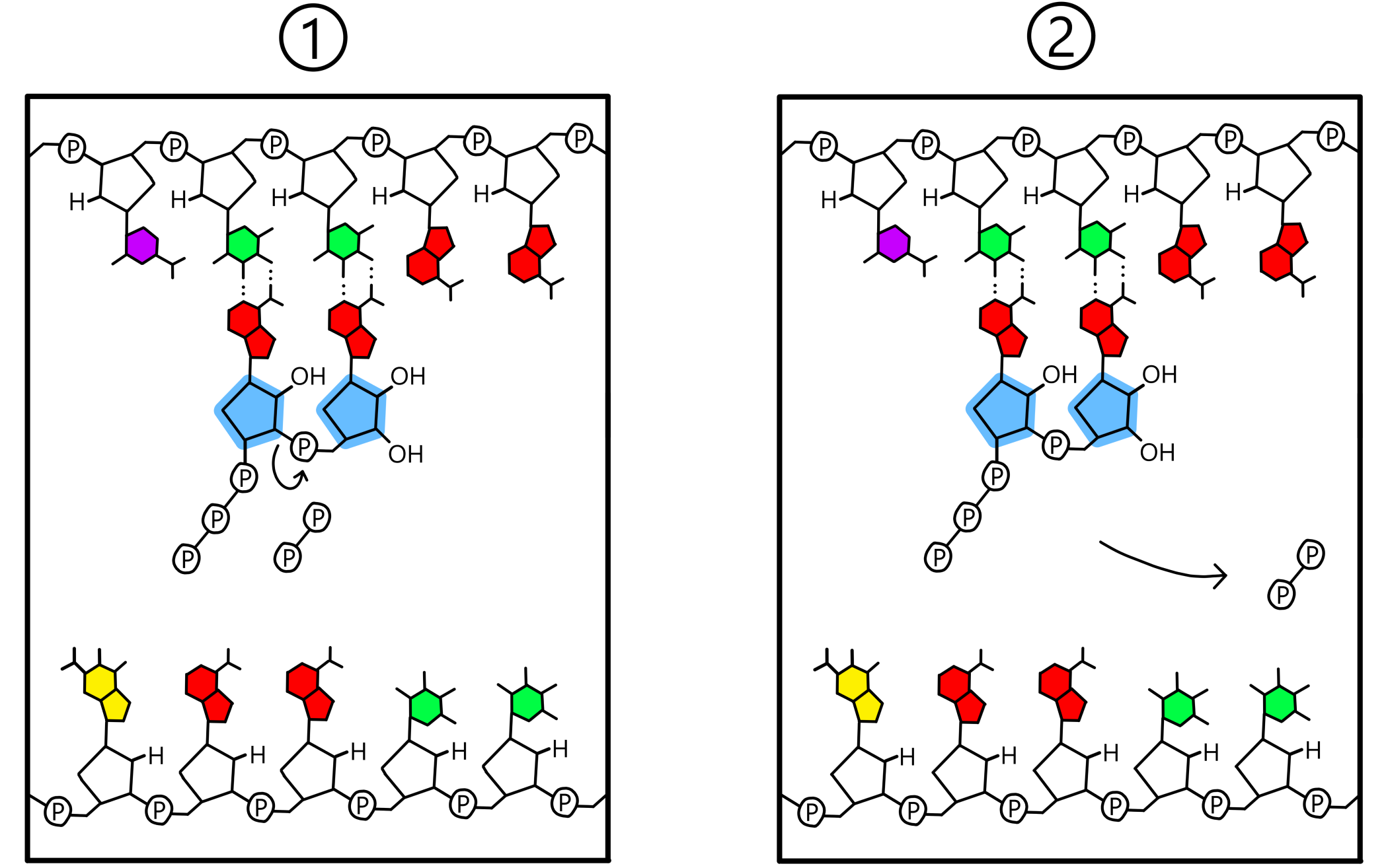
Når både P-stedet og A-stedet inneholder en nukleotid baseparet til templatet dannes det et kovalent bånd mellom de to nukleotidene. Dette båndet går mellom 3’-oksygen på nukleotiden i P-stedet og fosfat på nukleotiden i A-stedet. Templatet er nå bundet til en RNA-kjede bestående av to nukleotider (se Figur 3). (Ordet "RNA-kjede" er ikke helt passende enn så lenge bare to nukleotider har blitt inkorporert i kjeden, men jeg bruker det likevel.)
En detalj ved RNA-syntesen er at mens nukleotider i RNA har en fosfat hver, så bygges RNA fra nukleotider med tre fosfater hver. Hos disse såkalte trifosfatene er riboses 5’-karbon kovalent bundet til en første fosfat, som er kovalent bundet til en andre fosfat, som er kovalent bundet til en tredje. Det kovalente båndet som dannes mellom nukleotider går alltid mellom 3’-oksygen på nukleotiden i P-stedet og første fosfat på nukleotiden i A-stedet. Samtidig som at dette nye båndet dannes så brytes det eksisterende båndet mellom første og andre fosfat. Andre og tredje fosfat finnes deretter i form av en pyrofosfat, som løsner fra RNApol.
Etter bånddannelse mellom nukleotidene vil RNApol (samt A-stedet og P-stedet) bevege seg en distanse på en nukleotid langs templatet, i templatets 5'-til-3' retning. Dette gjør at ribose-nukleotiden i A-stedet flyttes over til P-stedet, mens ribose-nukleotiden i P-stedet flyttes ut av P-stedet. A-stedet er nå tomt og er posisjonert nær templatets tredje base. En ny ribose-nukleotid entrer A-stedet og danner et basepar med templatets tredje base, og igjen dannes et kovalent bånd mellom 3’-oksygen på nukleotiden i P-stedet og fosfat på nukleotiden i A-stedet. Templatet er nå bundet til en RNA-kjede bestående av tre nukleotider.
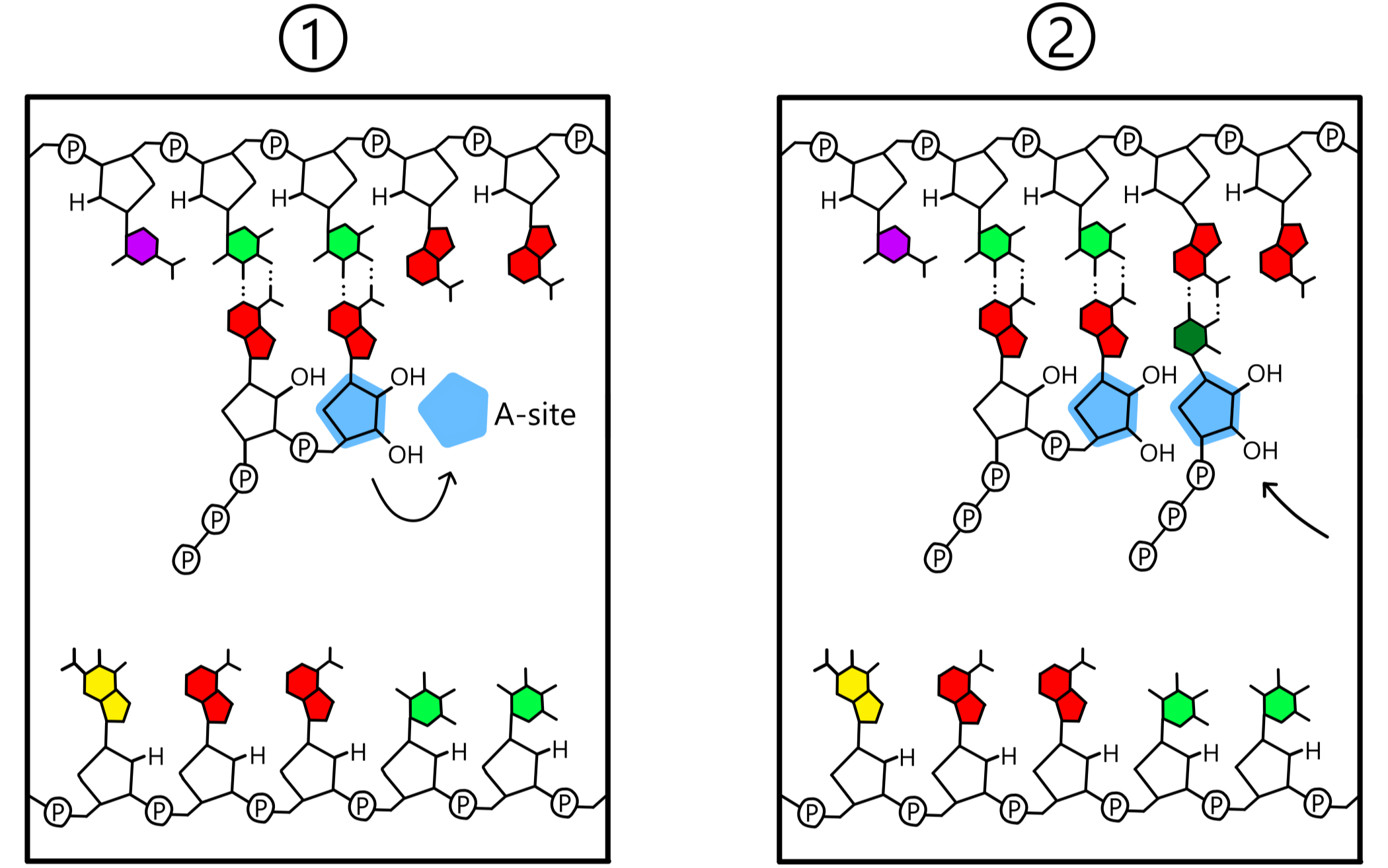
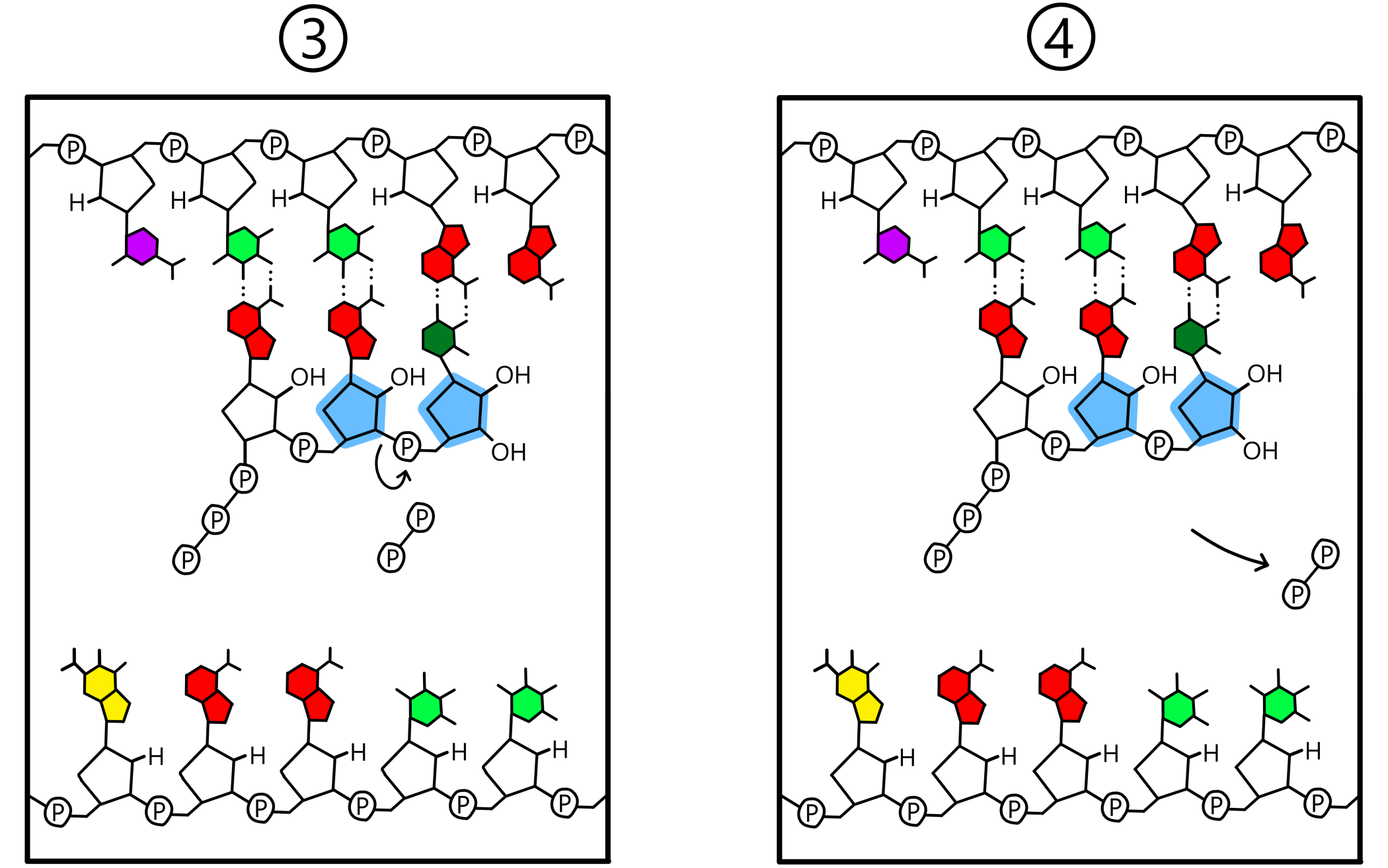
Hendelsene i forrige avsnitt gjentas om igjen og om igjen: RNApol beveger seg langs DNA, en ny ribose-nukleotid legges til RNA-kjeden, RNApol beveger seg, en ny nukleotid legges til, og så videre. Med dette vokser RNA lengre og lengre (se Figur 5).
Når RNApol beveger seg langs templatet vil de to DNA-kjedene i dobbelheliksen skille seg foran RNApol. Bak RNApol vil templatet skille seg fra RNA og binde seg til nontemplatet, slik at DNA-dobbelheliksen gjenoppstår. I praksis så vil transkripsjonsboblen bevege seg langs DNA sammen med RNApol. Dette betyr at så snart en RNApol har beveget seg bort fra en promoter så kan en annen RNApol binde samme promoter og begynne transkripsjon. Altså kan et gen transkripteres av flere RNApol samtidig.
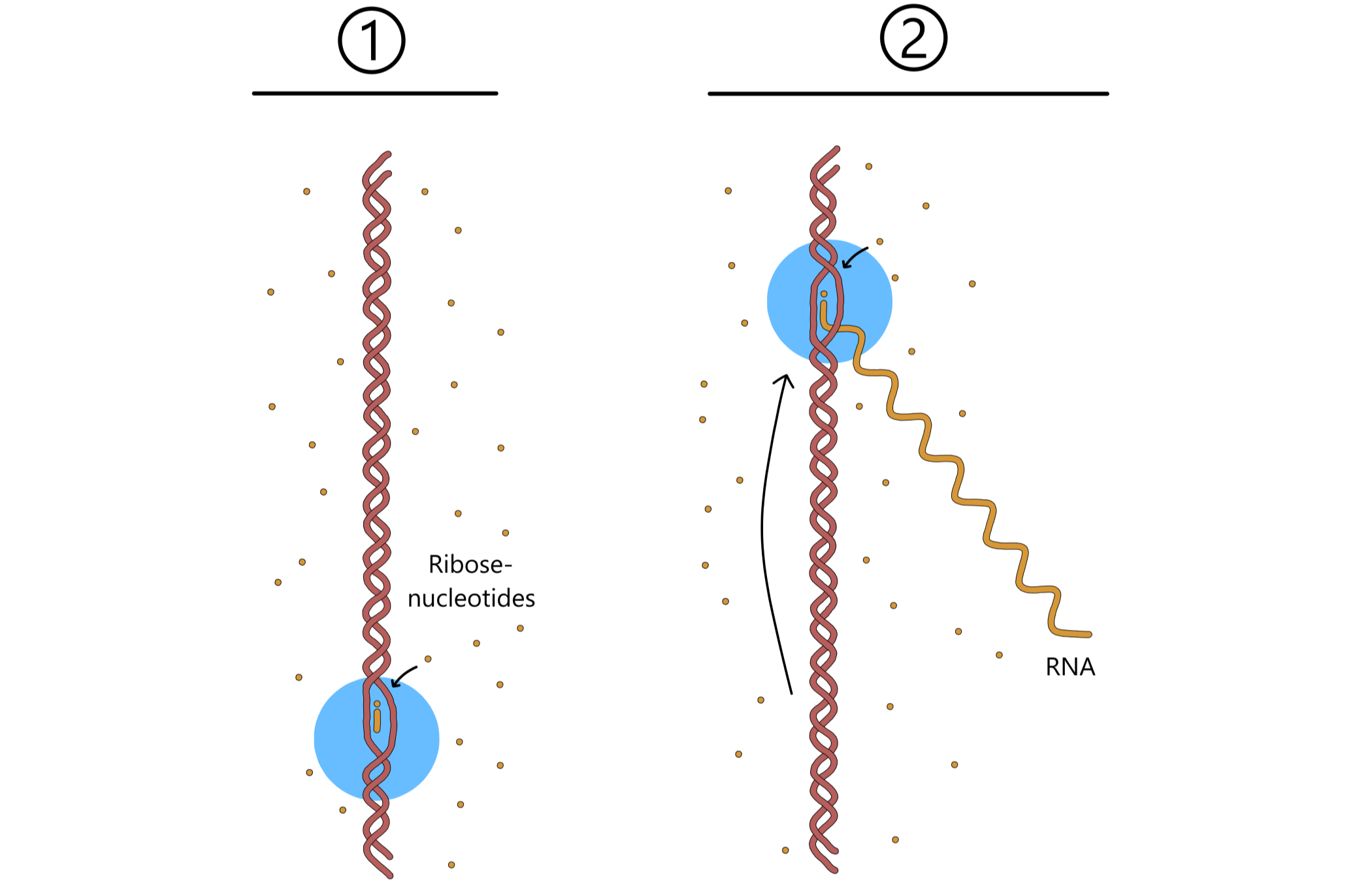
Eventuelt kommer RNApol til en terminator-sekvens i DNA. Denne sekvensen får RNApol til å stoppe opp. Deretter løsner RNA fra både RNApol og templatet, etterfulgt av løsning av RNApol fra DNA. De to skilte DNA binder sammen igjen slik at transkripsjonsboblen forsvinner, og med dette er transkripsjonen ferdig.
Merk at transkripsjon ikke begynner rett ved starten av et gen, og ender heller ikke rett ved slutten av et gen. I stedet begynner transkripsjon før genet og avsluttes etter. En RNA vil derfor ha en basesekvens ved hver ende som ikke oversettes til aminosyrer. På engelsk kalles disse delene av RNA for 5'- og 3' "untranslated regions".
Hvordan basesekvenser presenteres
Ved transkripsjonsstart kan nukleotider entre både P-stedet og A-stedet. Etter at RNApol har begynt å bevege seg vil derimot nye nukleotider kun entre A-stedet, ettersom P-stedet da alltid fylles av en nukleotid i RNA-kjeden. Det kovalente båndet dannes mellom 3’-oksygen på P-sted-nukleotiden og fosfat på A-sted-nukleotiden, så nye nukleotider vil alltid legges til ved 3’-enden av den voksende RNA-kjeden.
Tidligere ble det skrevet at transkripsjon er en slags kopiering av en basesekvens fra DNA til RNA. Sekvensen i RNA bygges antiparallelt og komplementært til sekvensen i templat-DNA, som igjen er antiparallell komplementær til sekvensen i nontemplat-DNA. Altså blir basesekvensen i RNA en nær kopi av basesekvensen i nontemplat-DNA.
Basesekvensen i RNA kan skrives på to ulike måter: i 3’-til-5’-retning eller i 5’-til-3’-retning. Siden RNA bygges i 5’-til-3’-retning, hvor nye ribose-nukleotider legges til ved 3'-enden av den eksisterende RNA-kjeden, så er det vanlig å skrive sekvenser i denne retningen (altså: venstre-mot-høyre skriveretning matcher 5'-til-3' sekvens-retning). Under DNA-replikasjon bygges DNA også i 5’-til-3’-retning, så det er også vanlig å skrive DNA-sekvenser i denne retningen.
Når geners basesekvens skal presenteres må en velge enten templat-sekvensen, eller nontemplat-sekvensen, eller begge to. Siden sekvensene i RNA og nontemplat nærmest er kopier så er det vanlig å presentere geners basesekvens i form av nontemplat-sekvensen. Promoter-sekvenser presenteres også i form av nontemplat-sekvensen, selv om promoter-DNA fungerer i dobbelheliks-form. Templat-sekvensen kan utelates da den er gitt implisitt av nontemplat-sekvensen ettersom de to sekvensene er komplementære. For eksempel, denne sekvensen fra dobbelheliks-DNA:
5’-TAATGTGAGTTAGCTCACTCAT-3’
3’-ATTACACTCAATCGAGTGAGTA-5’
Kan bare skrives som 5'-TAATGTGAGTTAGCTCACTCAT-3' (nontemplat). Dette er måten som vil brukes på denne nettsiden.
Ettersom RNA og DNA bygges i 5'-til-3'-retning så er det vanlig å referere til denne retningen som "nedstrøms". 3'-til-5'-retningen, som er motsatt retning, refereres til som "oppstrøms". For eksempel, dersom en sier at "På nontemplat-DNA er promoteren oppstrøms av genet", så betyr dette at promoteren er lokalisert i 3'-til-5'-retning i forhold til genet. (Uttrykkene oppstrøms og nedstrøms vil brukes en del i mine senere artikler, så de er verdt å huske om du har tenkt å fortsette å lese flere artikler.)
Sigmafaktorer
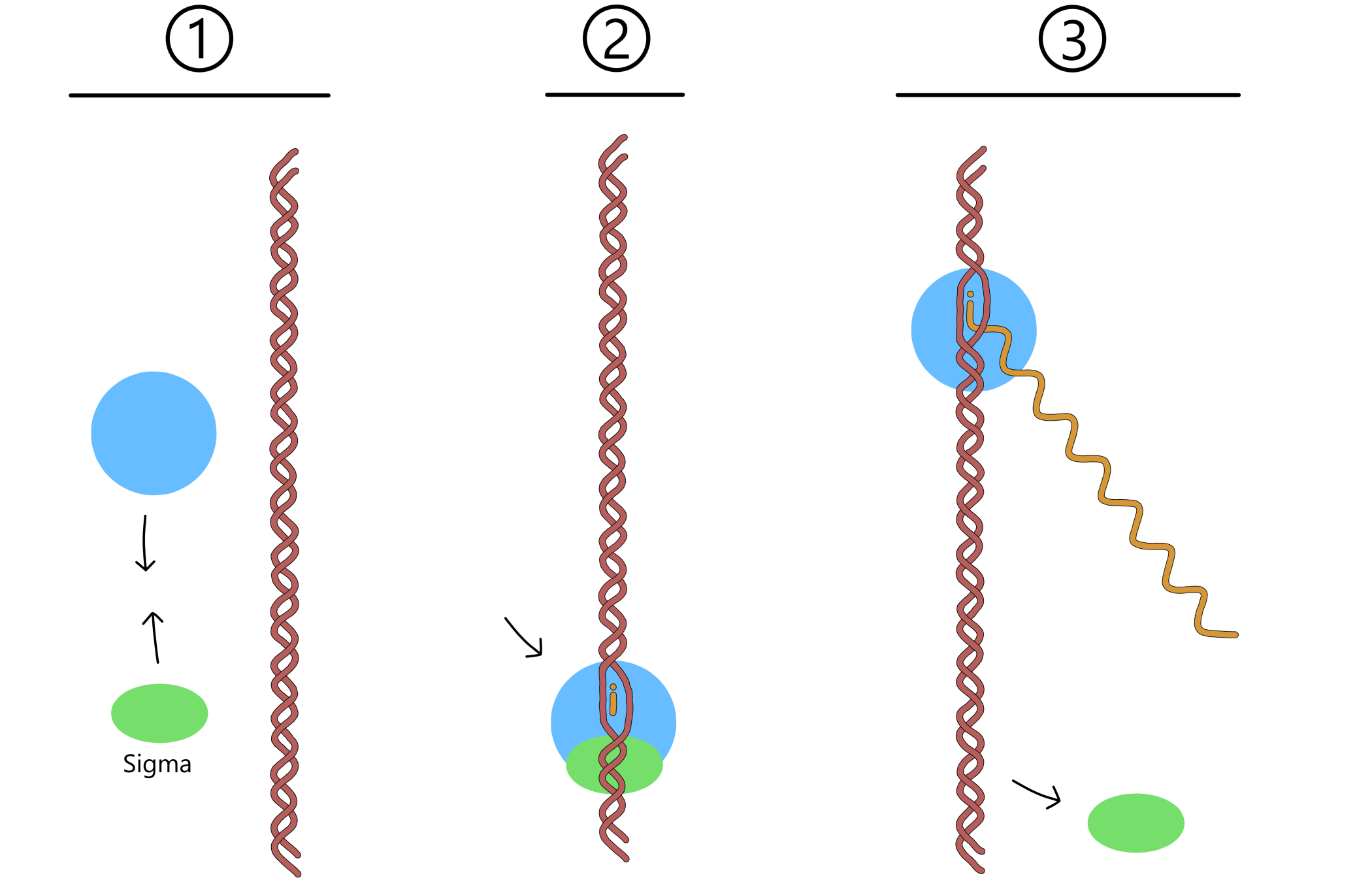
En viktig komponent i transkripsjon ble utelatt fra seksjonen om transkripsjonsprosessen, for å gjøre seksjonen litt enklere å forstå. Denne komponenten er et protein kalt sigma-faktor, eller bare sigma. Før transkripsjon kan begynne må RNApol binde sammen med sigma. Deretter kan sigma binde til en promoter, og RNApol binder da promoteren indirekte via sigma. Det finnes ulike typer sigma som lar RNApol binde til ulike typer promotere, i E. coli er det for eksempel syv ulike typer. Etter at RNApol har begynt å bevege seg langs DNA løsner sigma fra promoteren og fra RNApol, og tar ikke del i resten av transkripsjonen.
Oppsummering
Transkripsjon er en slags kopiering av en basesekvens fra nontemplat-DNA til RNA. Basene adenin, cytosin, og guanin kopieres eksakt, mens thymin i DNA kopieres til den lignende basen uracil i RNA.
Transkripsjon starter ved at RNApol binder til sigma, og sigma binder til en promoter-sekvens i DNA. Ved promoteren vil dobbelheliksen åpne seg til en transkripsjonsboble. To ribose-nukleotider entrer RNApol og baseparer med templat-DNA. RNApol danner så et kovalent bånd mellom den ene nukleotidens 3'-oksygen og den andre nukleotidens fosfat.
RNApol begynner så å bevege seg langs DNA. Nye ribose-nukleotider fortsetter å binde til templat-DNA, og RNApol fortsetter å danne kovalente bånd mellom nukleotidene. Med dette vokser RNA lengre og lengre. Eventuelt kommer RNApol til en terminator, hvor det stopper opp. RNApol, RNA og DNA løsner fra hverandre, og med det er transkripsjonen over.